Endosymbiondentheorie?
Fach: Biologie
Hat jemand eine Ahnung was mit Belegen bei dem Thema Endosymbiondentheorie gemeint sind?
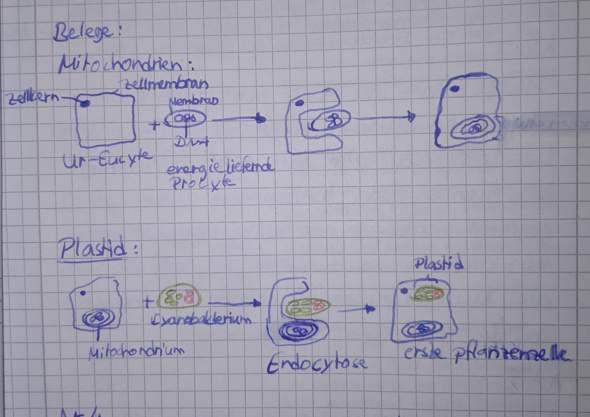
Ist das vielleicht das?
3 Antworten
Wie in deiner Zeichnung auch zumindest teilweise zu erkennen ist sind Mitochondrien und Plastiden in der Eukaryotenzelle von zwei Membranen umgeben (bei letzteren hast du die äußere in deiner Zeichnung vergessen). Die innere Membran ist eine ptokaryotische Membran (wie von Bakterien), die äußere eine eukaryotische. Daraus lässt sich schließen, das Mitochondrien/Plastiden ursprünglich Prokaryoten waren, die per Phagozytose von einer eukaryotischen Zelle aufgenommen wurden. Tatsächlich gibt es auch Belege für mehrfache Endosymbiose, also Fälle in denen dieser Eukaryot dann erneut phagozytiert wurde, dann befinden sich eben entsprechend noch mehr Membranen um die Plastiden.
Ein zweiter Beleg für die Endosymbiontentheorie ist die Existenz eigener ringförmiger DNA in den Mitochondrien/Plastiden, die ebenfalls auf ursprünglich eigenständige prokaryotische Organismen hindeutet.
Man merke in diesem Zusammenhang:
Eine wissenschaftliche Theorie ist anders als im gewöhnlichen Sprachgebrauch keine bloße Vermutung, sondern gewissermaßen der höchste Status, den eine wissenschaftliche Erkenntnis erreichen kann. Eine Hypothese wird erst dann zur Theorie, wenn sie von mehreren Seiten bewiesen und als wissenschaftlicher Konsens akzeptiert wurde.
Hi,
das was du da gezeichnet hast, ist die Vorstellung, die man davon hat, wie es abgelaufen sein könnte:
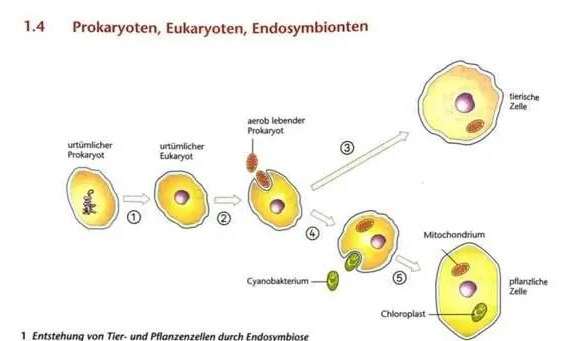
Belege sind zellbiologische Befunde, die diese Theorie stützen.
Dazu zählt z.B. die Beobachtung,
- dass die Organellen (Mitochondrien und Chloroplasten) sich niemals mit dem Zytoplasma vermischen, sondern durch eine doppelte Hüllmembran von diesem abgegrenzt sind
- dass Mitochondrien und Chloroplasten nur durch Teilung aus ihresgleichen hervorgehen
- dass Mitochondrien und Chloroplasten eigene, wie bei Bakterien üblich strukturierte, zirkuläre DNA, ohne kerntypische Histone oder Nukleosomen (kerntypische DNA-Verpackung) besitzen
- dass ihre Proteinbiosyntheseeinrichtungen bakterienähnlich sind, z.B. die Bauart ihrer Ribosomen, die sich von den Ribosomen der Zelle (des Zytoplasmas) unterscheiden
- dass sie ihr Erbgut eigenständig und von der S-Phase des Zellkerns (Zellzyklus der Zelle) unabhängig replizieren
Das sind Befunde, die darauf hindeuten, dass diese Organellen eine Sonderstellung in der Zelle einnehmen und Zellen, in denen sie vorkommen, sie nicht selbst entwickelt haben. LG
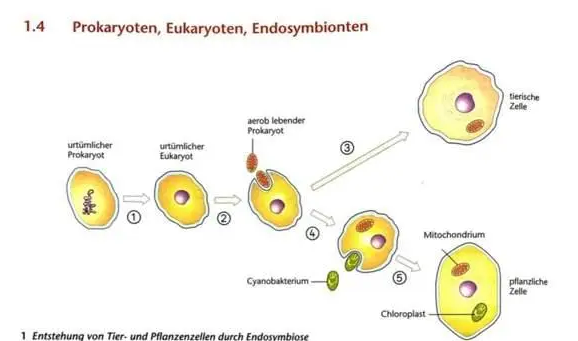
du hast das Modell widergegeben. Das Modell bietet jedoch keine Belege. Solche habe ich dir zusammengestellt. Gut, du hast oben in das Mitochondrium DNA reingezeichnet, das könnte man als ein Beleg durchgehen lassen :) falls du das auch so gesehen hättest. LG
Moin,
ja, deine Bilder haben damit zu tun. Es gibt eine ganze Reihe von Hinweisen auf die Richtigkeit der Endosymbionten-Theorie:
Gehen wir auf die Organellen Mitochondrium & Chloroplast ein...
– Das genetische System der Organellen hat prokaryotische Eigenschaften
- die DNA ist zirkulär
- der zirkulären DNA fehlen die typischen Histone von Eukaryoten-DNA
- die Transkription wird durch Rifamycine gehemmt (was einer antibiotischen Wirkung bei Prokaryoten entspricht); Rifamycine stören die Transkription von Genen bei Eukaryoten nicht
- die Ribosomen gehören dem prokaryotischen 70s-Typ an
- die Ribosomen sind Chloramphenicol-empfindlich (Antiobitumwirkung wie bei Bakterien), aber Cycloheximid-unempfindlich (das bei Eukaryoten die Translation hemmt)
– Die inneren Membranen der Mitochondrien oder Plastiden weichen von der typischen Eukaryoten-Membran ab
- innere Tiermitochondrienmembranen enthalten Cardiolipin (CL) (wie E. coli)
- sie enthalten kein Phosphatidylserin (PS) (ebenso wie es bei E. coli, Cyanophyceen oder Chloroplasten fehlt)
- Chloroplastenmembranen enthalten Phosphatidylglycerol (PG) (wie bei E. coli oder Cyanphyceen)
- außerdem enthalten sie Monogalactosylglycerol (MG), Digalactosyldiacetylglycerol (DGG) und Sulfoquinovosyldiacylglycerol (SDG) (wie Cyanophyceen)
– Das Fettsäuresynthese-System der Plastiden ist wie bei Prokaryoten
- es setzt sich aus einzelnen, isolierbaren Enzymen zusammen
- das eukaryotische System ist ein zusammenhängender Multienzymkomplex
– Indirekte Hinweise: Man kann heute bei unterschiedlichen Lebewesen verschiedene Stadien zwischen Symbiose und Endosymbiose beobachten:
- Korallen, einige Muscheln, der Wurm Convoluta roscoffensis, aber zum Beispiel auch Blattläuse leben in Symbiose mit Algen oder Bakterien, die im Zellinneren ihrer Wirte leben. Bei den endosymbiotischen Bakterien der Blattläuse werden beispielsweise Beschleunigungen der Evolutionsraten einhergehend mit Genverlusten und einem Anstieg des AT-Gehaltes der DNA beobachtet, wie sie auch bei Zellorganellen zu finden sind.
- Rezente Cytosymbiosen wie beim Pantoffeltierchen (Paramecium bursaria), das mehrere Hundert Zoochlorellen enthalten kann, ohne sie zu verdauen; der Wirt erhält von Chlorella-ähnlichen Symbionten Glucose, Maltose und Sauerstoff, der Symbiont erhält einen geschützten Raum, Kohlenstoffdioxid und anorganische Ionen.
- Der Süßwasserpolyp Hydra viridissima („Grüne Hydra”) kann durch Endocytose Zoochlorellen aufnehmen und mit deren Hilfe Fotosynthese betreiben.
- Manche Acoelomorpha, z.B. die Vertreter aus der Gattung Waminoa leben mit Zooxanthellen in Symbiose und ernähren sich unter anderem von deren Fotosyntheseprodukten.
- Die Arten der Convolutidae-Gattungen Convoluta und auch Symsagittifera leben in Symbiose mit der einzelligen Grünalge Tetraselmis convolutae zusammen.
- Ganz allgemein sind Zooxanthellen Protisten, die als Endosymbionten in einer Reihe von Lebewesen leben können.
- Die Lebensweise der Alge Haldena arenicola könnte als ein sehr frühes Stadium einer endosymbiontischen Beziehung zwischen zwei Arten angesehen werden.
- Die Wurzeln einiger Pflanzen leben in Symbiose mit stickstofffixierenden Bakterien (Rhizoiden; zum Beispiel Leguminosen).
- Bei Foraminiferen und Schwämmen kommen Rotalgen als Endosymbionten vor.
- Noctiluca scintillans (ein parasitisch lebender Dinoflagellat) nimmt Grünalgen der Spezies Pedinomonas noctilucae auf, die im Innern als Endosymbionten weiterleben.
- Bei anderen Dinoflagellaten sind verschiedene Stadien zu finden: Kleptoplastiden, komplexe Rhodoplasten und tertiäre Endosymbiosen, die auf die Aufnahme von Cryptophyceen oder auch Kalkalgen (Haptophyta), eine Gruppe mariner Algen zurückzuführen sind. Nachgewiesen wurde die tertiäre Endosymbiose zwischen Kalkalgen und Dinoflagellaten bei den Arten Gymnodinium breve, Gymnodinium galatheanum und Gyrodinium aureolum.
- Der Pilz Geosiphon pyriforme enthält endosymbiontisch lebende Cyanobakterien der Gattung Nostoc.
- Manche Blattläuse (z.B. die Erbsenblattlaus) können in besonderen Darmzellen (sogenannten Bakteriozyten) endosymbiontische Bakterien der Gattung Buchnera beherbergen, die über die Eier an die nächste Generation weitergegeben werden.
- In den Darmzellen von Motten-Schildläusen (Aleyrodoidea) findet man gleich zwei endosymbiontische Bakterien verschachtelt.
- Bei Schmierläusen der Spezies Planococcus citri wurde sogar eine verschachtelte „sekundäre“ Endosymbiose gefunden.
– Allgemeines:
- Plastiden und Mitochondrien sind von ihrem Aufbau her prokaryotenähnlich: kein Zellkern, ringförmige DNA; die DNA ist nicht mit Histonen assoziiert, sondern durch sogenannte HLPs verdichtet; Größe entspricht der von kleinen Bakterien.
- Plastiden und Mitochondrien stellen ihre eigenen Proteine her, wobei sie einen prokaryotischen Proteinbiosyntheseapparat besitzen. Ihre Ribosomen ähneln denen der Bakterien, nicht denen der Wirtszelle (≤ 70-S- anstatt 80-S-Ribosomen). Die mRNA der beiden Organellen besitzt nicht die für Eukaryoten typische 5'-Cap-Sequenz und auch die Polyadenylierung (Anheftung eines Poly-A-Schwanzes) fehlt.
- Die Cyanellen der Glaucophyta sind sogar noch von einer dünnen bakteriellen Zellwand umgeben!
- Rotalgen und Glaucophyta setzen wie Cyanobakterien Phycobiline zum Auffangen von Photonen in der Fotosynthese ein.
- Die DNA-Sequenzen der Mitochondrien ähneln denen der alpha-Proteobakterien, während Plastiden-DNA-Sequenzen im Cyanobakterien-Stammbaum platziert werden. Ein Vergleich mit der Wirts-DNA weist auf keine Abstammung der Organellen vom Wirt hin.
- Primäre Plastiden und Mitochondrien sind von Doppelmembranen umgeben, wobei, der Hypothese entsprechend, die äußere beim „Verschlucken“ des Bakteriums hinzugekommen ist. Die innere entspricht der von Bakterien (Vorkommen von Cardiolipin, kein Cholesterin; außerdem Vorkommen von Transmembranproteinen (β-barrel-Proteine), die nur in den Membranen von Bakterien und Zellorganellen vorkommen), die äußere entspricht der Membran von Eukaryoten.
- Die besten Belege für sekundäre Endosymbiosen finden sich bei den Chloraachniophyceen, zu den Cercozoa gehörenden Amöben und den Cryptophyceen, einer eigenständigen Algenklasse. Beide Algengruppen enthalten komplexe Plastiden mit vier(!) Hüllmembranen. Zwischen den beiden äußeren und den beiden inneren Hüllmembranen befindet sich der periplastidäre Raum mit einem Nucleomoprph, einem stark reduzierten eukaryotischen Zellkern mit je drei linearen kleinen Chromosomen und eukaryotischen 80-S-Ribosomen.
- Genomsequenzierungen und phylogenetische Analysen zeigten, dass Nucleomorph und Plastid der Chlorarachniophyceen auf eine sekundäre Endosymbiose mit einer Grünalge, der komplexe Plastid der Cryptophyceen jedoch auf eine sekundäre Endosymbiose mit einer Rotalge zurückzuführen sind. Von der Chlorarachniophycee Bigelowiella natans und von der Cryptophycee Guillardia theta wird/wurde das Nucleomorph-Genom vollständig durchsequenziert. Da bei Rotalgen die Stärkesynthese im Cytoplasma stattfindet und nicht wie bei den Grünalgen und Landpflanzen im Plastiden, spricht das Vorkommen von Stärke im periplastidären Raum der Cryptophyceen ebenfalls für eine sekundäre Endosymbiose.
- Mitochondrien und Plastiden vermehren sich durch Teilung und werden bei Teilung der Wirtszelle auf die Tochterzellen verteilt. Sie entstehen nicht de novo, das heißt, sie können von der Zelle bei zufälligem Verlust nicht neu gebildet werden. Und das bedeutet wiederum, dass ein Eukaryotenzellkern in seinem Genom keine Informationen zur Herstellung der Organellen besitzt.
- Die membrangebundenen ATPasen der Bakterien und Organellen (wie Mitochondrien) sind untereinander nahe verwandt, genauso wie die der Archaeen und die eukaryoteneigenen. Zwischen diesen beiden Gruppen besteht nur eine entferntere Verwandtschaft. Bei kleinen Gruppen von Bakterien und von Archaeen mit den „falschen“ ATPasen vermutet man horizontalen Gentransfer.
Du siehst, es gibt jede Menge gute bis sehr gute Gründe, die Endosymbiontentheorie für wahr zu halten...
LG von der Waterkant
Wow, eine der ausführlichstem Antworten, die ich auf GF bis dato gesehen habe.
Aber ist diese Antwort nicht vielleicht etwas Overkill? 😅
Der Fragesteller ist wahrscheinlich Mittelstufenschüler und wird mit vielen Begrifflichkeiten nicht viel anfangen können.
Ja, das befürchte ich in der Tat auch. Aber einerseits bin ich ein Fan von ausführlichen Antworten (sch... Perfektionismus) und andererseits mag ich den Song »Overkill« von Colin Hay (Sänger von Men at Work)... Was - so frage ich dich also - was sollte ich machen? Oder wie es Luther einst ausdrückte: »Hier stehe ich; ich kann nicht anders. Amen.« ;o))
Also ist das Falsch was ich gemacht habe?